
Access to all articles, new health classes, discounts in our store, and more!
Deficient Calcification Produced by Diet Experimental and Clinical Considerations
Published in Transactions of the American Therapeutic Society, 1939. From the Research Dept. of the Pottenger Sanatorium. Co-Author: D. G. Simonsen.
* * *
A normal supply of calcium and its utilization are important to an individual throughout life. Evidence is developing which indicates that a deficiency in calcium is not uncommon. It may be present during any period of life from birth to old age.
In order to influence calcification in patients, we have given diets rich in calcium, have administered quantities of calcium salts over long periods of time, and have administered vitamins to aid its assimilation; and yet we are unable to establish normal calcium utilization. Even though our ordinary diets are supposed to be adequate in the calcium content, it is only necessary to look at the teeth and facial development of people to see that something is or has been wrong with calcium metabolism in most of them.
Some factor or factors that are necessary for the assimilation and deposition of this mineral in the bones seem to be lacking in many people. Particularly may this be noted in children during the growing period of life. It does not seem reasonable to suppose that it is the mineral alone. Studies which we have made suggest that there are heat labile substances found in the dietary that exert control over the assimilation and deposition of calcium; and that these are partly if not wholly processed out of many of our foods by modern methods of preservation and preparation. We are not able to state their exact nature; but they appear to be substances which do not resist either heat or oxidation. In our experience we have found them to exist in high concentration in rapidly growing green grasses, in the raw flesh and glands of healthy animals, used as food. Such foods as milk and eggs of market grade, which are often recommended for their calcium content, are in our experience low in the important activating substances.
These factors cannot as yet be definitely classified with vitamins, hormones or enzymes. They may be any or all of them, or some entirely different factor, such as the physico-chemical state of the food. The lack or relative lack of this or these unstable factors produces a profound physiologic effect in many or all systems of the body. Though we are primarily discussing the calcification of bones in this paper, the nervous system, the respiratory, gastrointestinal and genitourinary tracts, the skin, muscular and endocrine systems, all show changes of importance when these stimulators of calcium metabolism are lacking.
Deficient Calcification Experimentally Produced
In our studies, the cat was chosen as the animal for experimentation. This discussion deals with the offspring of successive generations of animals on both sufficient and deficient diets. Our optimum diet when served to healthy cats from healthy parentage enables them to maintain themselves in excellent physiologic condition and produce healthy kittens. It consists of raw meat in which all parts of the animal are used, raw milk of market grade, and codliver oil. The deficient diet consists of the same three factors except that the meat is cooked. Limited experience suggests that cats do not do as well when pasteurized milk is substituted for raw milk in this diet. A high grade raw milk in which a large portion of the food fed to the cattle is obtained from green pasture or fresh cut green feed, will materially lessen the deficiencies caused by the use of cooked meat.
In our experimental work, calcification has been studied through several generations with reference to (1) kittens from healthy cats; (2) kittens from cats which showed evidence of thyroid deficiency; (3) kittens from cats which were in a lowered state of health from being fed on cooked meat; and (4) adult cats which have been placed on deficient diets after having had an opportunity to reach full development on adequate diets.
In analyzing the accumulated data shown in Tables 1, 2, 3, and 4, it is evident that the cats do not possess a constant degree of calcification throughout life. Neither do human beings, according to Todd.1 The ages of all cats analyzed in Table 1 were not known. In as much as some of the femurs may have come from cats younger than eighteen months when apparently the adult level of bone calcium is reached some of the figures for normal cats may be low. The age and entire life history of all deficient cats is fully known.
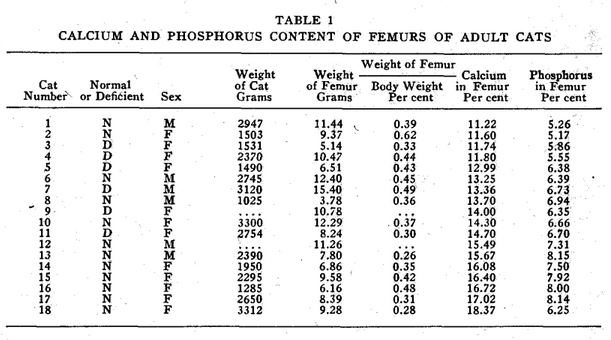
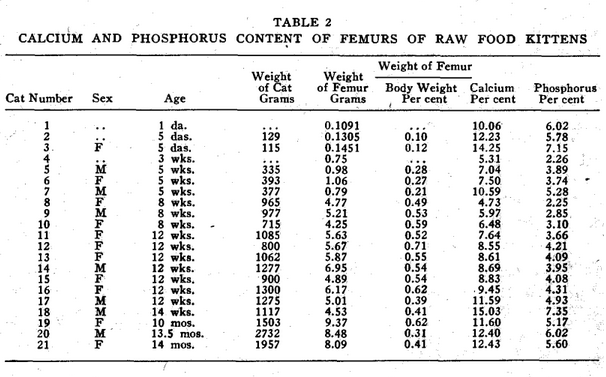
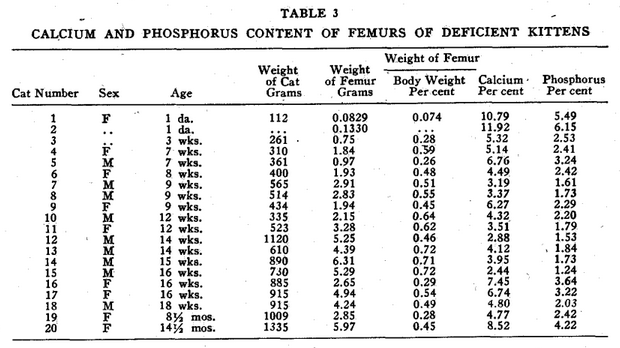
In our studies of cats, we confined our observations largely to the size and weight of the femur, and to the percentage of calcium and phosphorus which it contained.* In all groups studied, as shown in Table 4, the percentage of calcium and phosphorus at birth is practically the same as that of the mother cat. Shortly after birth, a drop in these elements takes place which is not restored as quickly in the deficient as in the healthy kittens. At the end of the first month, there is a second drop in these minerals which shows in normal kittens. This drop occurs about two weeks later in cooked meat kittens. In adult life, the calcium and phosphorus percentage of the bones of the cats from mothers on cooked meat remains lower than that of cats from mothers fed on raw meat. This difference was even more pronounced when there was evidence of thyroid deficiency in the mother.
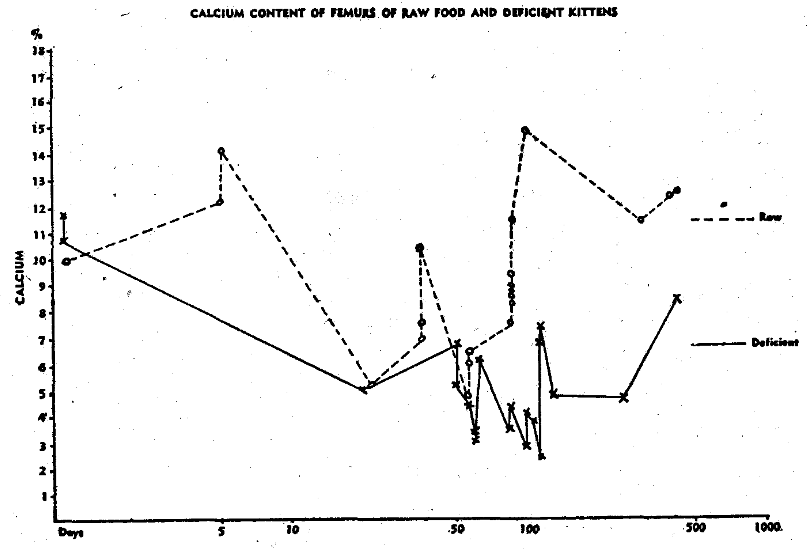
Fig. 1. Calcium content of femurs of raw food and deficient kittens.
Our diagnosis of thyroid deficiency in the female cat is based on (1) the physical characteristics of her kittens such as prominent frontal area of skull, small teeth, retracted lower jaw and failure of anterior development of the face; and (2) on pathological examination of the thyroid tissue upon her death.
In the cats which receive no raw meat, smallness of the diameter of the bones predominates.
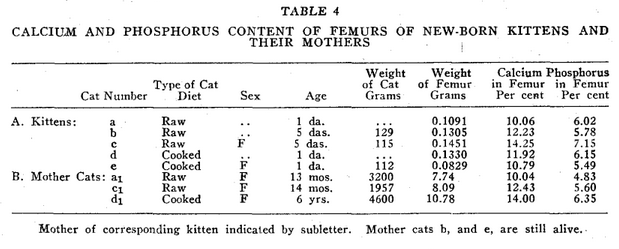
A typical healthy male cat from healthy parents, and maintained on raw meat, is shown in A, Fig. 2. He has a large skull, large bones, large thorax, large and long body, relatively short legs. His dental arches are broad. He has excellent teeth, regular and well spaced.

Fig. 2. Three tom cats eighteen months old. A, Healthy tom cat from raw meat parentage, maintained on raw meat all life. B, Deficient tom cat born from hypothyroid mother on raw meat. C, Cooked meat tom cat born of cooked meat mother.
The normal calcium for femurs of healthy adult cats of healthy parentage is from 12 to 17 per cent, and the phosphorus from 6 to 8½ per cent.
In cat B, Fig. 2, one sees a cat born of a mother with deficient thyroid, proved at autopsy. The mother had been on a raw food diet for a year preceding his birth; however, during the time she was nursing the previous litter, she had developed an abscess which was diagnosed by the character of some necrotic tissue obtained from the wound, as a thyroid abscess. This was later confirmed at autopsy. Though cat B was approximately the same age as cat A, it showed markedly inferior development. Typical cats from hypothyroid mothers such as described below, cat H, show a much lower percentage of calcium and phosphorus in their femurs. In cat B, there is a failure in the development of the face so that the teeth are crowded and narrow, the skull smaller than that of a well developed cat; the thorax smaller in diameter, the body shorter and the legs longer. In a similar manner all his soft tissues are inferior. The normal calcium of such femurs is 8 to 12 per cent and the phosphorus 4 to 6 per cent.
Cat C, Fig. 2, is an animal born of a cooked meat mother, and kept on cooked meat all its life. He has a small skull, very narrow dental arch, poor irregular dentition, small body and long legs. He is functionally sterile and all his soft tissues are of very inferior quality.
As an example of the bizarre conditions that can be produced by long use of cooked food and thyroid deficiency, the case of cat H is cited. She was a full sister of cat B of the previous litter. The mother cat had been on cooked meat for eighteen months, was placed on raw meat and bred immediately. As soon as she had delivered, she was put back on cooked food. Four kittens were born; two died within six weeks, one died in three months and the other in eight months. The report of cat H is herewith given in full. This mother developed a thyroid abscess during the nursing period, which was confirmed at autopsy. Because of its peculiar appearance the kitten was named “Streamline.” The legs showed marked bowing, the spine was distorted and the kitten developed a pot belly, which nearly reached the ground. It had a wizened appearance and when eight months old had only attained the stature of a six weeks-old kitten. About three months before death the cat showed signs of paralysis in the left hind leg, which increased in severity until all legs became involved; and one week before death the cat had several convulsive seizures. The weight at death was 1009 grams. Post-mortem examination revealed an under-developed animal with marked rickets, curvature of the spine and a rachitic rosary of the ribs. The bladder was enormous, measuring 2.5 x 3.5 x 1 inch, which contained 110 cc. of urine. This accounted for more than 10 per cent of the weight of the cat. The bladder had pushed the intestines to the right side of the abdominal cavity. The colon was greatly enlarged. The femurs were soft and spongy and contained almost no cortex. Analysis of one of them showed that it contained 4.77 per cent calcium and 2.42 per cent phosphorus. This was 60 per cent below normal for the age of the cat.
The report from the pathologist on cat H follows:
“Brain, cord and peripheral nerves show nothing of interest grossly. The bones are very soft and cut very easily. The long bones of the legs are markedly crooked. The cortex of the bones is thin and easily crushed. The marrow is abundant and quite fleshy.
Microscopic.–The heart shows nothing of interest. Two sections of lung show no evidence of inflammation or bronchitis. The bronchi have normal epithelium. Two small arterioles show gray thrombi. There is some hyperemia. Section of the trachea at the level of the thyroid shows normal findings. The epithelium is free from change. There is no thickening of the basement membrane of inflammatory infiltrate. The thyroid lobes included in the section show thyroid gland of an incompletely developed animal. The muscle about the thyroid is negative. Sections of the tubes and uterus show an exudate containing many polymorphonuclear leucocytes in the lumen but the mucosa is free from inflammatory change and is that of an immature animal. Sections of the stomach and intestines are free from noteworthy changes. The kidney and liver show simple cyanosis.
Sections of the muscles from the hind and front legs show considerable fatty infiltration of the muscles. The fibers are not atrophied. Many small nerve filaments in the muscles show no noteworthy changes. A few muscle fibers, however, in the piece show a fatty infiltration, have lost their striations, and stain more deeply with eosin than the other fibers. Sections were also made of many of the nerves of the brachial and lumbosacral plexuses, and most of these nerve fibers show normal architecture. The myelin is normally distributed in the fibers, but an occasional nerve shows much clumping of the myelin into irregular clumps. Some of the myelin stains deeper red with eosin than other portions, and there is disruption of some of the axis cylinders.
Sections through all levels of the cord, medulla and midbrain show no evidence of active inflammatory lesions in the meninges or in the cord or brain stem proper. The spinal nerve roots also appear free from noteworthy changes and numerous ganglion cells found in the posterior nerve roots are negative. The most striking finding is one that is found throughout the entire cord and brain stem, namely, a degeneration of some of the motor nerve ganglion cells, although some of the ganglion cells in parts of the posterior horns are also affected. Various degrees of change are seen, but most commonly one sees shrinkage and atrophy of the involved cells which stain very poorly and retain more eosin than the surrounding cells. Many are wrinkled or crumpled, their nuclei show various degrees of karyolysis, and the internal structure of the cells stains very badly. Some neuronophagia showing a typical collection of cells about the dying or dead cells is seen. Perivascular infiltrate or diffuse edema or hemorrhage, so characteristic of myelitis, is not present. All levels of the cord seem to be almost equally involved. No noteworthy changes in the white matter of the cord are demonstrated. The changes in the medulla involve the ganglion cells in the nerve centers in a fashion similar to that in the cord. Some degenerated ganglion cells are seen also in the cerebellum. These are fairly numerous. The cerebral cortex shows occasional degenerated ganglion cells, but little else of interest. The question arises whether the changes in these cells throughout the nervous system are merely post-mortem effects, but they do not have this appearance since the tissue appears to be well preserved. (Post mortem was performed immediately after death.)
The eye shows nothing of note.
Sections through many of the ends of the long bones show a marked disturbance of the histologic picture. In the femur and tibia, particularly, the line of ossification is narrow, the columns of cartilage cells are fairly regular, but the underlying osteoid and bone tissue is distinctly irregular and the trabeculae show active bone growth. There is no condensation seen in the normal bone and the shaft shows a very thin layer of trabeculated bone with fibrous marrow, rather than a thick cortical shell. Only occasionally is it fairly thick. In some places there is only the narrow zone of subperiosteal bone. In the heads of the bones the trabeculae are separated by a large amount of marrow. There is little reinforcement of the cartilaginous articular surface. The ribs show the same findings.
Diagnosis.–Osteomalacia–consistent with rachitic origin. Degeneration of ganglion cells in cord, brain stem, and cerebellum–etiology undetermined (no evidence of true inflammation). Slight degenerative changes in peripheral nerves. Fatty infiltration of some muscles.
The urinalysis report of this cat is as follows: Color, dark amber; turbidity, clear; specific gravity, 1.042; reaction, slightly acid; albumin, plus one; sugar, 0.63 per cent; acetone, 0; diacetic acid, 0; bile, negative; urobilin, negative; urobilinogen, trace at 1:10 dilution; casts, 0; leucocytes per H.P.F., 0-1; red cells per H.P.F., 20-30; squamous, numerous; round or oval, few; mucous shreds 0; amount of sediment, scant; and bacteria, occasional.”
Cat D (not illustrated) represents the fourth group of animals. He was born from healthy raw meat parentage, nursed by a healthy mother, kept for four months on raw certified milk, plus iron, manganese and copper. He had most excellent development. He was then placed on a cooked meat diet. After two and a half years, with the exception of having flabby tissues, he is still in excellent condition, and his skeletal description is practically that of cat A.
Figure 3 represents a comparative study of another group of kittens.

Fig. 3. E, Third generation cooked meat kitten twelve weeks of age. F, Second generation raw meat kitten from deficient mother. G, Second generation cooked meat kitten.
E, figure 3, is representative of a group of kittens which, so far, we have only been able to study up to the third month because we have been unable to get them to live longer. They are the most poorly developed and show bones most poorly calcified of all our groups.
In the illustration, cat E, figure 3, was a third generation cooked meat cat. His age was twelve weeks; his weight, 335 grams. His femur weighed 2.15 grams; calcium, 4.32 per cent; phosphorus, 2.20 per cent.
F, figure 3, was a second generation raw meat cat from a deficient mother cat which had been placed back on a raw meat diet. The cat was ten weeks of age; her weight, 523 grams. The femur weighed 3.28 grams; calcium, 3.51 per cent; phosphorus, 1.79 per cent.
Cat G, figure 3, was a second generation cooked meat cat. She was seven weeks old; her weight, 310 grams. Her femur weighed 1.84 grams; calcium, 5.14 per cent; phosphorus, 2.41 per cent.
Clinical Application of Experimental Data
An appreciation of the fact of the artificial production of calcium imbalance in these physiologically unstable kittens has enabled us to understand some of the observations which we have made on children in the clinic. We see many that seem to compare with these types. We have published a paper dealing with this phase of the subject elsewhere and would refer those interested in the subject to it.5
Figure 4 A represents an x-ray of a nine-months-old boy nursed nine months by a healthy mother subsisting on a diet rich in fresh foods grown on her own farm. He has bones of large diameter. His development age is normal, and he has fully recovered from the period of lowered calcification that follows birth. His skull is large, he has a well-developed dental arch and ample space for all his unerupted teeth. His soft tissues are of excellent tone. We have not been able to get bones from such a child for analysis, but x-ray studies suggest excellent calcification.
Figure 4 B represents an infant of similar age to A. The mother was a hypothyroid and unable to nurse the child. He was placed on an undiluted raw cow’s milk of the best grade obtainable. He has smaller bones, not quite as dense as those of A, but of fair quality. He shows a fair response from the normal lowered calcification following birth; has good facial development but inferior to that of A. Bones from x-ray studies suggest good calcification, but less than A. Tissue tone is good.
Figure 4 C shows the skeleton of an infant whose mother shows stigmata of deficiency. She was unable to nurse the child. He was placed on all kinds of cooked milk formulae. As a result, he made a poor recovery from the lowered calcification following birth. He shows delay in osseous development; has small bones; distorted facial structures, and is generally deficient in physiologic reaction.
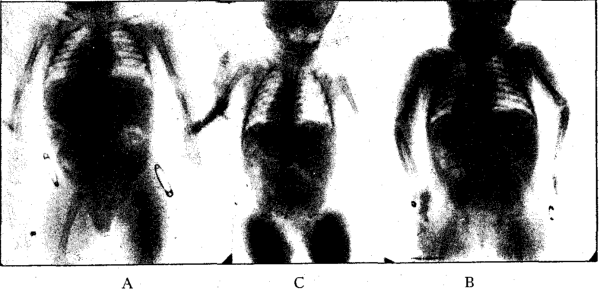
Fig. 4. X-ray of 3 nine-months-old infants. A, Healthy infant nursed by healthy mother. B, Caesarian infant born of hypothyroid mother, given raw cow’s milk formula. C, deficient infant from deficient mother; given cooked milk formulae.
Figure 5 shows the skull of a well-developed female, age thirty-eight, whose good development carried her through the growing period and until she entered upon adult life. Then, in order to “succeed” in her chosen work, she felt it necessary to economize, so she reduced her food intake. She lost her third molars and had cavities in three or four more teeth. Some of her bones are slightly more radiotranslucent than normal. Though suffering from a prolonged period of starvation, she still has an excellent body which has responded well to rehabilitation. Her height is 5 feet, 3 inches; she has large bones, a broad thorax, a large body and short legs. Upper and lower halves of body are of equal length.
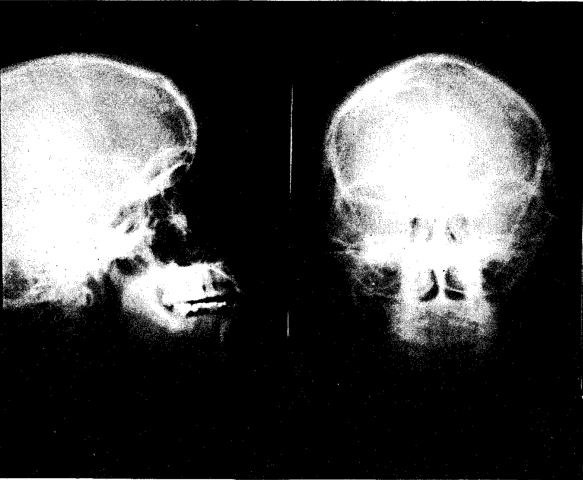
Fig. 5. X-ray of skull of female, age thirty-eight. Developed fully on good diet until age of nineteen, then on deficient diets for nineteen years.
Figure 6 represents the skull of an individual who belongs to the third generation of individuals on deficient diets. The family history of deficiency. includes the mother and grandmother. Her age is the same as that of Figure 5. She is 6 feet tall, and her bony structure shows poor calcification. She has small bones; a poorly developed skull; lost most of her teeth; small thorax; lower half of body much longer than the upper half. She has not had the vitality to work more than one day a week.
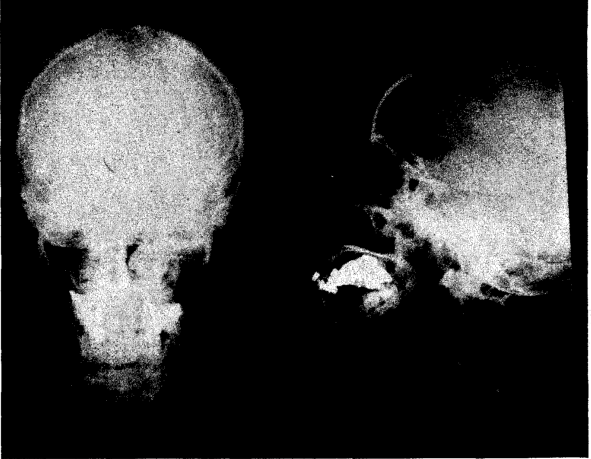
Fig. 6. Skull of female, age thirty-eight. Deficient development. History of deficiency in mother and grandmother.
It is our impression from our analysis of clinical and experimental data that several factors enter into the adequacy of an individual’s calcification. Unless the factors destroyed in deficient diets are added in excess to the dietary of individuals who are rendered deficient, little can be done to change the calcification that is established at the end of the child’s growing period.
This pattern of calcification depends largely upon the diet and health of the mother, but in this instance we have traced similar deficiency to the grandmother. It also depends on whether or not the infant was nursed; if not nursed, the type of formula used. This is well shown experimentally. This does not seem to be an inherited factor transmitted to the child by the germ plasm, but an acquired (environmental) factor passed to the growing embryo by the chemistry of the mother. The maternal influence, which we are describing here, depends upon the mother’s diet and health and the diet and health of her mother, and judging from our experimental work probably that of more remote generations. We have experimental evidence to show that four or more generations may be necessary to restore a strain of cats to normal after they have been rendered deficient by diets, so that they can bear litters of normal kittens.
An adequate amount of these heat labile factors is necessary in the mother’s milk as well as in substitute foods in order to bring about the proper calcification of the skeleton. These factors are largely destroyed in pasteurized and boiled milk formulae. For the growth and development of a child nothing can replace the milk of a healthy mother fed on an adequate diet.
An important physiologic factor in determining the bone development of the infant and later of the adult is the potency of the maternal thyroid. A disturbance in thyroid function in the mother and, or, her ancestors shows in the development of the offspring.
When animals have had excellent calcification through their growing period and up to the time of the closure of the epiphyses of the long bones, they have an excellent start in life. Those who are poorly developed and show low calcification during the early years of life are less able to stand the usual stresses to which they are subjected during life.
Discussion
We have described some deficiencies produced in cats by cooked food diets. In this discussion, we are summarizing experiments that have been continued over a seven-year period. We are primarily discussing the deficiency of successive generations of offspring of animals rendered deficient by dietary means. It is particularly important to know that under the conditions of the experiment which must be recognized as being far more rigid than those applying to human beings, that the deficiency renders the experimental animal so deplete in important vitalizing factors that the third generation is unable to live beyond the period corresponding to childhood in the human being.
In our experimental work, we have confined our observations to an animal that is normally carnivorous. Man has adapted himself both to herbivorous and carnivorous diets, but we feel that the experimental evidence gained from the study of a truly carnivorous animal is applicable in either instance. The human infant, according to modern conception, should be nursed for a period of nine months. During this period of life, the normal human being, like all other mammals, subsists upon a food substance of animal origin. Further, anthropological studies have shown that the best developed human beings are heavy meat eaters. The human infant should receive his nourishment direct from the source without any heat processing. On the other hand, primitive man who has assumed a carnivorous diet consumes a considerable portion of his dietary uncooked. In a like manner, those people who are largely vegetarians in their dietary habits, have consumed a great deal of fresh fruits and vegetables and rapidly growing sprouts in which heat labile factors are not destroyed.
In applying the facts which we have learned from our laboratory studies to the clinics we have been able to improve the physiologic response of children who have previously been developing in a deficient manner similar to the animals we have discussed.
In our clinical work we have used the highest grade of raw milk, raw meat, raw vegetables and fruit products obtainable. We have found that the extensive use of meats, milk, eggs, fresh fruits, vegetables and sprouts of highest quality in the raw state aid children with such deficiencies as are herein described to attain a more normal development.
Footnote:
*Calcium and phosphorus were determined on whole bone. Upon death of the animal, the femurs were removed and placed in an ice chest at -25°C. for 24 hours. At the end of this time, all fat and muscular tissue could easily be peeled off the bones. Organic matter was destroyed by oxidation with nitric and perchloric acids.2 Calcium was determined by titration with permanganate, a modification of the method used for serum calcium being used.3 Phosphorus determinations were carried out by a modification of the method of Fiske and Subbarow.4 The amounts of calcium and phosphorus found are expressed in terms of per cent of the whole femur.
References Cited:
- Todd, T. Wingate: Atlas of Skeletal Maturation. St. Louis, The C. V. Mosby Co., p. 42, 1937.
- Gerritz, H. W.: J. Indust. & Eng. Chem. Anal., 7:167, 1935.
- Practical Physiological Chemistry. Hawk & Bergein, ed. 2, p. 408, 1926.
- Fiske, C. H. and Subbarow, Y.: J. Biol. Chem., 66:375, 1925.
- Pottenger, F. M., Jr., and Pottenger, F. M.: “Adrenal Cortex in Treating Childhood Asthma: Clinical Evaluation of Its Use.” Calif. and West. Med., vol. 49 (Oct.), 1938.